
Abstract
A new fossil trackway is described in the upper lacustrine Miocene in the Prebetic Zone of the Iberian Peninsula, in Jumilla town (Murcia region) called Aenigmatipodus jumillensis nov. ichnogen. nov. ichnosp. This trackway consists of a pattern made up of sets of three tracks or triads, which are subparallel to each other, arranged in alternate groups. Each track presents a depression formed by a central body that is three times as long as it is wide, with straight or slightly curved walls, with two shorter bodies placed at the ends, one of the ends being shorter and more pronounced than the opposite, which is longer and stretched. All the biomechanical possibilities compatible with an anatomical design that could leave the impression of three alternate triads of tracks are analysed. The supports are only from the extremities on one side of the organism (left or right), the displacement being by translation. It is concluded that it had to be a large arthropod (metre scale), with a hexapod or decapod (less probably octopod), which had to be dragged laterally by a current in a very shallow lake or wetland environment. To date, no fossil organism is known, nor its current equivalent, that corresponds to these characteristics.
Resumen
Se describe una nueva pista fósil en el Mioceno superior lacustre de la Zona Prebética de la Península Ibérica, en la localidad de Jumilla (Región de Murcia) denominada Aenigmatipodus jumillensis nov. ichnogen. nov. ichnosp. Esta pista presenta un patrón que está formado por conjuntos de tres huellas o tríadas, subparalelas entre sí, y dispuestas en grupos alternos. Cada huella presenta una depresión formada por un cuerpo central que es tres veces más largo que ancho, con las paredes rectas o ligeramente curvadas. Presenta también dos cuerpos más cortos situados en los extremos, siendo uno de ellos más corto y pronunciado que el opuesto, que es más largo y estirado. Se analizan todas las posibilidades biomecánicas compatibles con un diseño anatómico que pudiera dejar la impresión de tres apéndices alternos. Los apoyos son sólo de las extremidades de un lado del organismo (izquierda o derecha), siendo el desplazamiento por traslación. Se concluye que tuvo que tratarse de un artrópodo de gran tamaño (escala métrica), ya fuera un hexápodo o decápodo (menos probablemente octópodo), que tuvo que ser arrastrado lateralmente por una corriente en un entorno lacustre o humedal muy poco profundo. Hasta la fecha, no se conoce ningún organismo fósil, ni su equivalente actual, que responda a estas características.
Similar content being viewed by others


1 Introduction
Sierra de las Cabras is a remarkable site due to the great variety of fossil tracks from the upper Miocene. Herrero et al., (2022, 2023) have already recognised and described the tracks of eight vertebrate ichnogenera (seven mammals and one bird). This work aims to add to the site the description and study of the tracks of a giant invertebrate not known until now. These tracks form a relatively long rake made up of an association of three tracks repeated at least seven times. The tracks and associated structures were studied, as well as the possibilities of displacement according to the spatial arrangement of the tracks in the rake. As a result, it has been deduced that the tracemaker is a hexapod arthropod that moved laterally, supported only by the three limbs on one side. Both the form of movement and the identification and size of the tracemaker, which is more significant than some of the mammals that cohabited with it in the environment, are noteworthy.
2 Geographical and geological location
As already mentioned, the new ichnofossil was found in the Sierra de las Cabras tracksite (hereinafter CBR, a simplified abbreviation to refer to the Sierra de las Cabras). This site is located eight kilometers westwards from the town of Jumilla (Murcia Province, SE Spain) at the southern foothills of the Sierra de las Cabras (Lat. 38º 28′ 53″, Lon. 1º 24′ 52″, datum: ETRS89; Fig. 1). From a geological point of view, the Sierra consists of folded and faulted Jurassic and Miocene carbonate rocks area belongs to the Prebetic Zone (outermost part of the External Zones of the Betic Cordillera). Tectonic contraction mainly occurred during the middle to late Miocene times, in the late orogenic stage of the Betic orogen (e.g., Roca et al., 2006; Rubinat et al., 2013). Then, the Prebetic Zone experienced drastic paleogeographic changes associated to the tectonic closure of the so-called Betic Strait, a marine seaway that connected the Atlantic and the Mediterranean up to the early Tortonian (Martín et al., 2009). In the time frame of this large-scale geological process, the shallow marine systems with extensive carbonate sedimentation that had prevailed during early Langhian to early Tortonian times gave way to a complex landscape of intramountain basins where marine sedimentation was rapidly replaced by continental deposits (e.g., Rossi et al., 2015).

Geographic and geological location of Sierra de las Cabras site and geographic location of other Mio-Pliocene tracks sites of SE Iberian Peninsula
The tracks beds considered in this paper are stratigraphically located in a bedding plane (Figs. 2, 3) at the top of a centimetric-scale limestone bed stratigraphically located in a ~ 10 m thick stratigraphic unit consisting of thinly bedded, fine-grained limestones and marls. It unconformably overlies Serravallian marine deposits and is in turn overlaid by post-orogenic yellowish marls of Pliocene age that cover large areas of the foothills of Sierra de las Cabras (Baena, 1981). In two recent papers (Herrero et al., 2022, 2023) assigned to this unit a latest Tortonian to earliest Messinian age, based on stratigraphic criteria and regional correlation.

The Sierra de las Cabras (CBR) site. a Diagram of the site with the study surfaces and the tracks. b The cleaned track ensembles. c Enlargement with the CBR192 trackway and intersecting vertebrate footprints

General view of the holotype CBR192 trackway
Sedimentary facies of the Sierra de las Cabras site are dominated by cm-scale bedded fine-grained often chalky limestones and some marly beds which correspond to a variety of freshwater, very shallow lacustrine, and palustrine carbonates. These have been interpreted as deposited in ponds, swamps, and marshes that should characterize a complex wetland system in which freshwater was probably provided by springs that could have formed at the foot of the mountain range, within the highly porous Jurassic and Miocene carbonate rocks (Herrero et al., 2022). The local availability of freshwater given by those wetlands could attract the diverse fauna reflected by the rich ichnofossil record of the site (Herrero et al., 2022, 2023).
Shallow freshwater deposits are mostly represented by micritic facies that contain abundant ostracod shells and small gastropods (Hydrobia sp., usually less than 3 mm high). The presence of calcified green algae and cyanobacteria is remarkable, forming small bioherms dominated by calcareous tubes possibly attributable to the genus Cladophorites, but a variety of microbial forms can be recognized. Palustrine facies show pedogenetic features such as micro-nodulization, circumgranular-cracking, clotted fabric, pedogenic-ooids, root voids, and Microcodium, pointing to dominantly wet environments in swamp—marsh settings. On the contrary, desiccation cracks, evaporite molds and evidence of early dolomitization processes suggest drier conditions with negative hydric balance that could prevail during relatively long periods of time. These climate considerations are concordant with regional climate trends reconstructed for that time interval in southern Iberia, characterized by increasing continentality, seasonality, and aridity (e.g., Dam, 2006; Jiménez-Moreno et al., 2010) although possibly punctuated by at least one wetter period that occurred around 7.0 Ma (De Miguel et al., 2019).
3 Palaeoichnology
The method used in the study of the present tracks was the same as the one carried out in the same outcrop by Herrero et al., (2022, 2023).
The images were processed with Adobe Photoshop® and translated on a scaled plane using AutoCAD. The dimension and orientation data are measured directly on the outcrop and on the AutoCAD representation. For 3D processing of the images Adobe Photoshop®, Agisoft Metashape Professional 1.5.1.7618 software, MeshLab®, and Paraview® software were used.
3.1 Ichnotaxonomy
Ichnogenus Aenigmatipodus nov. ichnogen.
3.1.1 Name derivation
From the Latin aenigma, enigma, a reality that cannot be understood, or that can hardly be understood or interpreted, and from the Greek ίχνος, which means trace.
Aenigmatipodus jumillensis nov. ichnoesp.

Subunit (even track) CBR192.7z. a Natural track. b Image with significant lines. c Diagram with the significant lines and parts (α, β, γ) of the print. d Image with contour lines and depth in false color. The scale is 2 × 10 cm

Subunits and indirect structures associated with the tracks. a Even track CBR192. 5 × rectangular shape of the boundary line of the central body β. b Even track CBR192.3z steps shape of the front part of the central body; β possible sole of the appendage with grooves that continue at the trailing end; γ oblique burrs from mud extrusion. c Odd track CBR192.4x. d Even track CBR192.5z, steps at the edge of the central body (β) in the vicinity of the inlet end (α) and grooves at the outlet end (γ). e1 Odd track CBR192.4z. Striations at the end of β and at the trailing end (γ), e2 detail of the box. f1 Collapse of the wall of CBR192.6y; f2 drawing of the structures; f3 contour lines of the track. g CBR192.2y deformed by CBR.111.1 footprint (Ursipeda). h CBR192.4z deforming to CBR101.4p-m (Canipeda). Scales: sheet, 2 × 10 cm; lines at (g and h), 10 cm
3.1.2 Diagnosis
The trackway is formed by a series of tracks that are grouped in sets of three tracks or triads, each of them being subparallel to each other and arranged in alternating sets. Each track constitutes a subunit of the whole set and consists of a depression or cleft formed by a central body and two bodies placed at the ends. The central body is approximately three times as long as is wide, with straight or slightly curved walls and longer than the two ends. One end is shorter and steeper, while the opposite end is longer and stretched.
3.1.3 Type material
The trackway constitutes an iconotype (sensu Bromley et al., 2003) since the material is not collectible. The holotype (CBR 192) and subunits are preserved in situ; however, a field number has been assigned to each of the subunits studied: CBR 192.1x, y, z; CBR192.2x, y, z; CBR192.3x, z (Fig. 5b); CBR192.4x (Fig. 5c), z; CBR192.5x (Fig. 5a), y, z (Fig. 5d); CBR192.6x, y, z; CBR192.7x (Fig. 4), y, z.
3.1.4 Type locality
Sierra de las Cabras, Jumilla (Murcia).
3.1.5 Type horizon
Upper Miocene (latest Tortonian-earliest Messinian).
3.1.6 Name derivation
From the town of Jumilla, where the outcrop is located.
3.1.7 Material
Nineteen tracks distributed into sets or triads that each have three subparallel tracks: CBR192.1, CBR192.2, CBR192.5, CBR192.6 and CBR192.7, and two sets with two tracks: CBR192.3 and CBR192.4.
Type ichnospecies Aenigmatipodus jumillensis nov. ichnoesp.
3.1.8 Diagnosis
Aenigmatipodus presents the shorter end of each track with oblique striations in the direction of the central body; the longest end is more complex, almost always with mud piled up towards the outside of the track, presenting grooves and fine, straight or slightly curved grooves at the bottom; the central body is a rectangular, deep hollow, with vertical side walls and a flat bottom. All triads are the same orientation of the three parts of the tracks (central body and two ends). All tracks have the same orientation of polarity.
3.1.9 Description
The tracks occupy an elongated sector with direction N12E at level 3 of the site. They form a sequence of seven repetitions of triads of parallel tracks (Figs. 2, 3, and 6). Although in the first set (CBR192.1) there are five tracks and in others (CBR192.3 and CBR192.4) there are only two, in the rest (CBR192.2, CBR192.5, CBR192.6, and CBR192.7) there are three. We interpret that in CBR192.1 there may be overlapping of tracks, or perhaps some of the marks are not related to the trackway, and that in CBR192.3 and CBR192.4 the three tracks are not preserved or have not been printed. The width of the trackway is about 70 cm and its length is 325 cm.

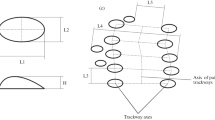
Schematic of the trackway CBR192. a Tracks and position of the midline of the triads with the designation (x, y, z) of each of its three tracks. b Drawing of the axis of the tracks with the direction of expulsion of the mud. c Position from the z track center, of the: f line that joints the center point of z and x tracks; d parallel line to perpendicular lines to β axis; and g parallel line to perpendicular lines to middle point to γ exit directions. We consider the line (P) that joins the center of the straight lines f step (or stride) and the line that joins the center of the successive P lines we consider the median line (ml)
To discern each of the three tracks we name them x, y, z from West to East, for example, the triad CBR192.7 contains the tracks CBR192.7x, CBR192.7y, CBR192.7z. The tracks consist of a central body (β) and two ends (α, γ) (Figs. 4c, 7). Almost all tracks are asymmetrical in the sense that their drawing is a sinuous object with the ends pointing like an S or a Z. The orientation and placement of the three parts of the tracks are the same in each triad: the ends α (North end) and γ (South end); and the central body β between the two previous ones (in the center of the track). Taking the central body axis β as a reference, the striations at the α end of the three tracks (x, y, z) of a triad point to the same side, while those at the γ end point in the opposite direction. In the even tracks, the γ end points to the SE, while in the odd ones, it points to the SW. Thus, tracks CBR192.1, CBR192.3, CBR192.5, and CBR192.7 would be the even ones and CBR192.2, CBR192.4, and CBR192.6 would be the odd ones. The three segments are subparallel in the tracks of each triad.

Longitudinal section and parts of the tracks of CBR192: (α) entrance, (β) central body and (γ) exit end. 1 and 2 are the points where the depth of the tracks has been measured
The central body (β) is generally the longest. It is a depression or hollow, whose outline is sometimes a rectangle (CBR192.7x, CBR192.7z, CBR192.5x, CBR192.5z) that has vertical walls (Figs. 4, 5a). The mean dimensions (Table 1) of the central body are (track length) l = 11 cm (maximum//minimum = 19//5 cm), (track width) a≈ 4 cm (maximum//minimum = 6//3 cm). The average depth of the central body is about 2.6 cm [2.7 cm at point 1 and 2.5 cm at point 2 (Fig. 7)]. In many tracks, the surrounding rim is deformed either because it has sunk slightly (possibly due to gravitational fall from the walls) or, on the contrary, because it is somewhat raised (in the form of a mud extrusion rim). The sole of the central body is not preserved in any of the tracks, except perhaps in CBR192.3z (Fig. 5b). However, it does not present irregularities, so it was probably flat. The proximal zone of the central body β or the zone of union between β and end α, can be rounded, occupied by a landing, or by small steps (Figs. 4, 5a–d). Taking into account the dynamics of the movement of the supports and the position of the extremities in the body, it follows that the proximal part of the supported feet must be located at the junction between the β and α bodies. At the junction with the γ, there are grooves beginning at the sole of β (Figs. 4, 5b). Both the grooves and the steps are repeated in many tracks.
The α end is shorter than the central body, and sometimes depressed. It has striations (Fig. 4) that look like drag striations. The direction of the grooves is oblique to the direction of elongation of the central body.
The γ end is more complex and often has mounds of mud towards the outside of the track (Figs. 4, 5e1–2). On the bottom, there are fine, straight, or slightly curved striae and grooves that can continue over the piled-up mud (Figs. 4, 5c–f3). The mean length of the γ end is 9.5 cm (maximum//minimum = 12//4 cm). The angle α^γ formed by the ends is a little more than 160º (or a little less than 20º if the supplementary angle is measured).
3.1.10 Remarks
We consider that all the mentioned structures (except those of fall of the walls, mud extrusion burrs, and mud piles) are direct structures (Gatesy, 2003) formed in the contact between the surface of the appendage with the mud. Tracks are real tracks in the sense of Requeta et al. (2006–2007) and Sarjeant (1989). We do not know if there is any stamp among them (Brown, 1999), nor therefore the structure of the appendage because we do not know well the base or the sole of the central body (β), because it is not distinguishable or because, when it is somewhat distinguished, it is occupied by striations or grooves (i.e., deformation structures). The mud dragged from inside the track, together with the superimposed striations, indicate that the γ end belongs to the end of the sequence of events for the generation of the tracks, that is, to the K phase of Thulborn and Wade (1989). The morphology of the appendage is erased in γ by the inherent drag at its exit; it is possible that the millimeter-sized grooves and the finer striations are produced by skin projections of the foot and by pilosities.
In line with the above, the striations at the end α must be prior to the entry of the appendage into the track, that is, from the initial phase of the track or phase T. The steps at the front of β, in the vicinity of α, are static structures that are possibly due to the support of skin or exoskeleton with wrinkles or scales.
The straight, vertical sidewalls (β) imply that the sides of the appendage were straight and relatively tall. These were probably made of dermal tissue that did not deform when penetrating the mud (very hard or chitinous skin). The steps, the vertical walls, and the formation of the hole are associated with the W phase.
The track sequence can be examined under two different considerations: that the triads are tracks of the limbs of one side (left or right) or that they are tracks of the limbs of both sides (left and right). In both cases, it is necessary to deduce which part is the one printed by the anterior (proximal) or posterior (distal) sector of the foot.
Apparently, there is a difference between the ends of the appendage so that one can think of its polarity. In what follows we will start from the hypothesis that: (i) the difference in impressions (steps versus grooves and striae) and; (ii) the direction of movement of the tip of the appendage implies that α–β is the proximal part of the track and β–γ the distal part. This determines that: either all the traces of the trackway are tracks from both sides; or if they are only of the same side, we would have to deduce if they are left or right.
Other indirect structures have been mentioned in the previous section (mud extrusion rims, deformation produced by the fall of the walls, and possibly, the mud accumulated at the γ end. Of these, the gravitational fall of the wall leads to the collapse of the track as in CBR192.6y (Fig. 5f1–3). Other indirect structures are found such as interferences with vertebrate footprints. This is the case (Figs. 2, 5g) of an ursid footprint (CBR11. 1) that pushes the mud that twists and narrows CBR192.2y, or in the case in which the tracks of CBR192 deform those of another trackway with which they intersect (Fig. 5h). These last two examples allow us to establish that the author of CBR192 left its tracks in the same time interval as the rest of the vertebrates on surface 3.
The genesis of the CBR192 tracks is synthesized in the following phases:
-
Passage of vertebrates (Ursipeda and possibly others).
-
Phase T, formation of α striae. Phase W, formation of the track hollow with the proximal steps, the sole and walls of the hole, and the extrusion rims. Phase K, grooves in the β–γ zone, accumulation of mud and grooves and striae in γ; fall of the walls of the track.
-
Passage of vertebrates (Canipeda and possibly others).
4 The trackway
The ensemble of all the triads (Fig. 3) is ordered in the same way as a trackway formed by the theoretical sequence of three tracks; the trackmaker would have to be a hexapod arthropod, like insects, or else, if it had more appendages (i.e., a decapod arthropod) it would only leave the mark of three of them.
The orientation of the three bodies (α, β, γ) of the tracks of the even triads is the opposite of that of the odd triads (in the same way as the letters “S” and “Z”). All the tracks have the same polarity orientation (α to the North, γ to the South. Figs. 6 and 7).
To visualize the relationships between the triads of the trackway, a line (f) is drawn in each triad that joins the central point of x–z, another (d) parallel to the perpendicular lines to the β axis, and another (g) parallel to the perpendicular lines to the axis of γ. We mark the midpoint of line f as a reference point to draw and measure P, a dimension that we will consider the pace length. We consider the line that joins the center of the successive paces to be the median line (ml) of the trackway (Fig. 6).
Line f oscillates east and west along the trackway with what appears to be a to-and-fro motion. If we place the origin of lines d, f, g in the middle of the central body (β) of the alternating tracks (x–z) of the end of the triads (Fig. 6) it can be observed that:
-
Line f forms an open angle towards the West, consistent with the arc formed by the median line. In general, the x track of each triad is placed further north than the other two.
-
Line d of a triad is subparallel to the line g of the two adjacent triads and vice versa, or what is the same, the axis of the central bodies β of a triad (for example, CBR192 6), is subparallel to the axis from the γ end of the adjacent triads (CBR192.5 and CBR192.7).
-
The pace length (P) and the pace angle (Ap) measurements of the trackway are similar (Table 1).
4.1 The support of the appendages
The movement in and out of the pes appendage when forming the track is reflected in the ends of the print (α, γ). The greater pressure of the appendage on the ground is responsible for the formation of the central body (β). Movement is also indicated by the orientation of the grooves, such that the direction of entry and exit are similar and the appendage runs SE in the odd tracks (and triads) and SW in the even tracks. There is no criterion that indicates rotational movement of the appendage during its entry into the mud, that is to say, it is possible that it descended vertically once contact with the ground was established.
We assume that the appendages of the three tracks of each triad move at the same time and that all three tracks are imprinted at the same time. The joints of the three extremities (slip-in, maximum weight support, drag-out) do the same action during the genesis of each triad. The sequence of triads implies a sequence of movements.
4.1.1 Support sequence
On the other hand, and as discussed below, the previous arguments indicate that the organism moved either laterally (in which case the triads can be from one or both sides of the arthropod's body), or longitudinally if the triads are from alternate sides. If it is assumed that the producing organism could be an insect, they only move (except in exceptional cases) in the direction marked by the alignment of the triads, which does not agree with the tracks observed at the site. This aspect is discussed later in the section related to the possible trackmakers of these tracks. In the case of hexapod arthropods with movement similar to eurypterids (Briggs & Rolfe, 1983), the placement of the tracks would indicate movement toward the North.
The analysis of the sequence of supports of the tracks has several interpretations that depend on several assumptions:
-
The first refers to the polarity of the appendages, that is if the morphology of the tracks depends: (i) on the formation dynamics (the appendages are symmetrical with respect to a transversal axis, Fig. 8b, c) or; (ii) on the structure of the appendage with the proximal part different from the distal;
-
The second, if the individual supports the extremities on one side or on both sides (left and right extremities) and
-
The third, if the movement back and forth of the limbs is significant with respect to the possible to-and-fro movement of the trackmaker’s body [limbs on the same side (see Sect. 4.2, below)] or if the position of the parasagittal axis rotates towards left and right from one triad to the next (Fig. 9a, b).

a Trackways with steps and midline according to the interpretation of groups of successive tracks; lateral displacement, transversal to the anteroposterior axis of the models. b Model of a thin-bodied trackmaker. c Model of a thick-bodied trackmaker. The width and length of the trackmaker’s body are random

Longitudinal displacement. a Limbs under the body; Position model of a thick-bodied trackmaker in each of the supports. b Position of the parasagittal axis relative to lines. d, f, g. c Relative position of the tracks and the axis E. d Position of the tracks for a sequence of supports if the arthropod moves with the anteroposterior axis along a straight line; the triads are oriented with the same tendency as in other hexapods (see Briggs & Rolfe, 1983; Getty et al., 2017). e Model of a hexapod with lateral supports; the body of the hexapod is minimally separated from the track
In the event that the triads have alternating sides (left and right), the only movement allowed is with the parasagittal axis oriented parallel to the elongation of β, that is, perpendicular to line d (Fig. 9a, b).
4.1.2 Supports with longitudinal displacement
There are examples of hexapod arthropods (Briggs & Rolfe, 1983; Getty et al., 2017) in which the parasagittal axis of the body is parallel or tangential to the midline, and in which the placement of the triads is symmetric with respect to that axis; we will call it longitudinal displacement. If we consider that the parasagittal axis of the arthropod is as indicated (approximately parallel to the body β or perpendicular to the line d, Fig. 9c, d) the position of the triads of the CBR192 track may be concordant (Fig. 9d) with the scheme presented by Briggs and Rolfe (1983) and by Getty et al. (2017). The orientation and placement of the triads with respect to the trajectory can be interpreted as congruent with the models of the previous authors if the body of the arthropod rotates between one support and the next (Fig. 9); this can be appreciated if we place the parasagittal axis of the model on a rectilinear trajectory (Fig. 9d).
4.1.3 Supports in lateral movement
We consider that the parasagittal axis or axial line of the trackmaker should be parallel to line f (Figs. 6, 8), which also implies that all three limbs have the same length or a similar functional length given the aligned placement of tracks x, y, z (Fig. 8a–c). The exit of the appendage from the ground can obey two types of action. Thus, the mound of mud at the γ end can be due to the fact that during the K phase the appendage is pushing the trackmaker’s body (active appendage), or because the appendage is dragged by the body (passive appendage). Both possibilities will imply completely opposite effects: lateral displacement towards the East or towards the West and progression towards the North or towards the South. If the displacement is active, the progression is towards the North and that of the body from West to East in the transition from the even to the odd triad (for example, from CBR192.6 to CBR192.7), while in passive displacement the progression is towards the South and the movement of the body is from East to West in the transition from the even to the odd triad (for example, from CBR192.6 to CBR192.5). The sequence of supports of the triads depends on the process of accumulation of mud in the γ body.
If the triads are on both sides (left and right), the arthropod's body position is incompatible with the footfall sequence, as shown below. Furthermore, if the animal supports both extremities, the movement that determines the position of the dragged mud is rotation around a vertical axis that is not shown on the track.
4.2 Compatibility of possible models
According to what has been said, it must be considered whether the trace of tracks is formed: first, by the support of the appendages on both sides of the body [left and right, [Figs. 9, 10, 11, 12)]; or second, only by the support of the appendages on one side [left or right (Figs. 13, 14, 15)].

Support of the two extremities (left and right). a By turn. b By lateral displacement. c Support in CBR192.4-CBR192.5 –red- and in CBR192.6-CBR192.7 –blue-. (Also the supports could have been in CBR192.3-CBR192.4 and in CBR192.5-CBR192.6). d For support to occur, one of the extremities must be directed forward and the other backward

a–c The left and right limbs are supported at the same time. On each support, two even triads (blue-violet) or two odd triads (red-green) are stepped on. d For support to occur, some limbs must be directed forward and others backward

a–c Alternate support of the appendages on one side (red-green) and on the other side (blue-violet) by twist. The contact of the sole of the appendage with the ground depends on the movement capacity of its joint with the rest of the extremity. d For support to occur, the extremities must be at the same height

a–c Unilateral limb support (blue to green) with reciprocating front-to-back movements. Supports in the CBR192.6 and CBR192.7 triads. d Movement possibilities of the trackmaker according to the length of the extremities and the rotation of the joint of the extremity with the body

Alternate limb support and body movement. a, b The body moves in the same way as line f. c, d The body moves in the opposite direction to line f

Example of the variability in the progression of a trackmaker with relatively long limbs and joints with a wide twist. a, c Northward progression – appendages (limbs) active (drive) in phase K. b, d Southward progression- appendages (limbs) passive in phase K. Crossed red stripes in c and d represent that the two constructions are impossible
If the appendages are supported on both sides, two possibilities must also be taken into account: if the parasagittal axis of the arthropod coincides or is tangent to the midline of the trackway, or if it is totally oblique to it. In the first case (longitudinal displacement) the body of the arthropod must rotate around a vertical axis (Fig. 9a–c, e); in the second, [lateral displacement, (Figs. 10, 11, 12)] it can be by rotation of the body (the axial plane rotates 360º every two supports around the parasagittal axis of the body) or by translation with the horizontal body [the axial plane of the body is vertical (Figs. 10, 11)] or with the body tilted [the axial plane makes a certain angle with the tracking surface (Fig. 12)].
In longitudinal displacement (Fig. 9), with the parasagittal axis of the animal's body parallel to the orientation of β, the direction of advance is towards the North due to the position of the tracks (cf. Briggs & Rolfe, 1983) and due to the placement of the mud expelled in phase K. Sector α would be the distal part of the track. If we take the orientation of the parasagittal axis parallel to β (perpendicular to line d), said orientation changes from one triad to the next, oscillating around an axis that is vertical to the animal's body. This type of displacement has several drawbacks:
-
The lack of intermediate groove marking and the little separation of the triads implies that the animal’s limbs must be below the body.
-
The depth of the three tracks is the same, so the position of the body was probably horizontal.
-
The appendages have to move outwards at each step so as not to interfere between the successive triads; given the great length of the limbs and the short length of the step, the movement they must make between one support and the next, cannot be justified without incompatible movements of the limbs and body of the animal.
The feet penetrate the mud vertically, and there are no structures similar to swimming marks, but consistent support on the bottom. If the extremities are placed under the body and their proximal joint is lateral, it is inferred that they must be greater than the separation between the internal and external tracks (x and y), that is, more than 70 cm. If the limbs were under the body of the animal but placed laterally, the rotation of the body and the position of the tracks between each two supports does not help the movement, but rather complicates it excessively (Fig. 9e). The model is incompatible.
In lateral displacement, if it is considered that the trackway is formed by the support of the left and right appendages (Figs. 10, 11, 12), it must be accepted, as has already been said, that these are proximally-distally symmetric. In this case, the difference in the shape of the tracks is not due to the difference in the shape of the appendages, since the proximal (internal) part of one triad has the same shape as the distal (external) part of the next—the α part of the tracks is distal in the triad on one side, and proximal on the other (unlike the γ part, which would be proximal in a triad and distal in the adjacent one). In this case, there are several possibilities that we differentiate.
-
The first, is that the even triads are from the extremities on one side (Figs. 10, 12) and the odd ones on the other. In this case, it is also necessary to distinguish between two types of progression, depending on whether the supports are by rotation (Figs. 9a, 12) or by translation (jump, Fig. 10b).
-
The second (Fig. 11) is that the even triads are from the left and right extremity (from a support) and the odd triads from the left and right extremity (from the adjacent support). Successive supports are by jump.
-
If it is considered that the trackway is formed by the support of the appendages on both sides (left and/or right), in displacement by rotation two options had to be considered
-
The first (Fig. 10) in which the axial plane between two successive supports has to rotate 360º and the support is at the same time alternate extremities (left–right);
-
The second (Fig. 12) in which the axial plane between two successive supports has to rotate 180º if there is only one triad (left or right) resting on the ground.
If it is considered that the trace of the tracks is formed by the support of the appendages on one side [left or right (Fig. 13)], the axial plane is always inclined towards the same side.
Finally, it remains to examine the movement of the body (axial line E) with respect to that of line f. The axial line E may not move, maybe in the same direction as that of line f or in the opposite direction (Fig. 14). These possibilities depend on the functional length of the limbs and the movement of their joints.
4.3 The progression
If we consider: (i) the displacement is lateral; (ii) the triads are always on the same side; (iii) the position of the mud expulsion zone, (iv) the placement of the extremities and the body, two displacement hypotheses can be considered (Fig. 15):
-
First, the mud is expelled by the force of the end of the appendage that propels the body of the trackmaker. The progression is towards the north (Fig. 15a).
-
Second, the mud is expelled by force from the end of the appendage, driven by the movement of the trackmaker’s body. The progression is towards the south (Fig. 15b).
The position of the appendages does not depend on that of the body, but on the joints, which can move independently with respect to the body of the trackmaker. It cannot be deduced, therefore, if it advanced towards the north or towards the south, at least if the extremities were long enough.
In summary, the positions in lateral displacement that imply simultaneous support of the two triads (six limbs) (Figs. 10, 11) have several drawbacks:
(i) the polarity of the tracks; (ii) the non-symmetrical position of the limbs; (iii) the opposite movement of the feet on one side and the other in phases T and K, (iv) the distance between the tracks on the support and between two successive supports.
Also, in the case of alternating support of the extremities on each side, the movement by rotation of the animal’s body around its axial axis is incompatible (Fig. 12) due to the polarity of the tracks and the variation in the speed of rotation necessary to maintain the equidistance (P) between the triads.
As a conclusion, the supports are only from the extremities on one side of the organism (left or right), the displacement being by translation. The following are compatible: (i) the polarity of the tracks; (ii) the symmetrical position of the extremities; (iii) the movement of the feet during the T and K phases of the footfall, and; (iv) the distance between the triads.
However, it must be specified that:
-
i.
In this model, the sway of line f, may or may not condition that of line E; the westward and eastward swing of the axial line depends on the length of the limbs and the magnitude of their movement back and forth, parallel to the axial line.
-
ii.
The progression towards the South of the organism occurs with passive displacement and can be independent of the length of the extremities.
-
iii.
Northward progression can only be justified if the tips of the appendages propel the body of the organism during exit and the limbs are relatively long.
In none of the cases analyzed can we deduce the anterior or posterior position of the ends of the axial line, that is, towards where the possible hexapod had its head.
Both types of progression imply that the animal is subjected to a lateral force that maintains the inclination of the body. If we assume that the proximal part of the track is the one placed to the North of the triads, the body’s belly must be placed to the north of the triads, with which, in both passive and active displacement, the force is directed to the South, opposing it. It is feasible to suppose that this force comes from a current of water that goes towards the South, probably of little intensity, given the relatively large volume and the probably small weight of the animals that made the tracks.
A summary of the possibilities described above is shown in Table 2.
5 Interpretation. Possible producing organisms
Due to the previously described characteristics, the tracks are directly compatible with an organism that had six appendages arranged in a series of three on each side of the body. The only hexapod organisms compatible with this organization are insects.
Other options could be those of organisms that have more appendages on each side, which would only leave the impression of three. This is the case of decapods (five pairs), or some octopods, such as scorpions or spiders (four pairs).
To date, no fossil organism is known, nor its current equivalent, that corresponds to these characteristics.
5.1 Size considerations
In the first case, the determination of the size of the possible insect producing these tracks can only be estimated approximately from the distance between the separation of the first (track x) and the third of each triad (track z). This separation has an average value of 55 cm, which implies that we would be, a priori, facing one of the largest insects in the fossil record.
Currently, the largest insects are among the Coleopters (beetles) such as Batocera wallacei (Wallace’s long-horn beetle), which can reach 26.67 cm in length, Dynastes hercules (Hercules beetle) of 18.1 cm and Titanus giganteus (Titan beetle), 16.51 cm. (Wood, 1983).
In the case of Batocera, the length of the body is approximately 75% of the separation between the anterior and posterior appendages. In Dynastes, that length is 115% greater than the distance between appendages, and in Titanus, the length is 95% of that separation. If we applied these proportions, the possible insect could have a variable length, between 41 and 63 cm.
Other of the largest are some blatodeos (cockroaches). The largest fossil cockroach found is the Apthoroblattina, a giant cockroach from the Carboniferous, with species that could measure between 43 cm (Bolton, 1921) and 45 cm in length (Bolton, 1911), and which was almost 3 times larger than the largest living cockroaches (Megaloblatta blaberoides) found in South and Central America, and can reach up to 18.5 cm (Walker, 1871). In this case, the length of the body is approximately 130% greater than the separation between the appendages, which, extrapolated to our case, would give us a length of the trackmaker of around 71 cm.
5.2 The lateral movement
However, apart from the size, a crucial aspect is related to its movement. All the biomechanical possibilities compatible with track registration have been exposed, where basically there are two types:
-
i.
The model in which the organism would move the extremities on one side that would be displaced (forward or backward) with respect to those of the other (backward or forward) in relation to the axial line. The organism may rotate, or not, with respect to this line, moving with the aerial phase, and
-
ii.
The model in which the extremities are supported alternately or only on one side, without the possibility of turning the body.
In both cases, the displacement is lateral, alternate, with a zigzagging movement of the axial line of the body. This type of movement is not compatible with a forward movement of a live insect, such as beetles or cockroaches make. Both move with a gait called “alternating tripod,” where the hind or forelimb on one side moves simultaneously with the middle limb on the other side of the body. This means that there are always at least three limbs touching the ground, which improves its stability. Or, as happens with some beetles, the movement is “galloping jumps” taking impulse with the middle extremities, leaning on the front ones, and dragging the rear ones.
This type of movement would leave marks that would be oriented antero-posteriorly in the direction of advance of the organism, arranged one after the other, following a row or being slightly displaced between them, a pattern that does not correspond to what was observed at the site.
On the other hand, only very few insects, such as some Zygentoma of the Lepismatidae family (Lepisma saccharina, Thermobia domestica), have the ability to move laterally. However, their size (about one centimeter) and their organization are very different (they have two long antennae on the head, three appendages under the back of the head, which are similar on the back, a long caudal filament, a last segment of the abdomen and two cerci), which makes it impossible for it to be the possible trackmaker.
5.3 Appendages and their relative sizes
Furthermore, the tracks of the triads are of similar sizes, which implies that the limbs that produce them should also be of a similar size. And this character is also against a beetle-cockroach type organization. These insects have disparate limb sizes. Those in the anterior position (prothoracic) are the shortest, those in the middle position (mesothoracic) are intermediate in size, and those located in the posterior position (metothoracic) are the longest. For all these reasons, the most feasible movement option is that of a body that was dragged laterally. And the most favorable agent responsible for this type of displacement would be a water current, perhaps driven by the action of the wind.
The fact that the marks present grooves of millimeter dimension at the end and finer grooves produced by projections of the appendage and possibly by pilosities, could indicate a hexapod with a swimming habit with appendages adapted for this purpose. This is the case of the great diving beetle (Dytiscus marginalis, Linnaeus, 1758), a species of adephagous beetle of the Dytiscidae family that lives in bodies of fresh water, although it needs to come to the surface to breathe (but its size is 2.7–3 0.5 cm in length).
The appendages of the possible hexapod should be quite long (the length of the legs with the feet parallel would have to be large), which could also indicate a swimming habit, as in the case of the current Notonecta (an aquatic insect belonging to the family Notonectidae, although they measure from 0.8 to 1.6 cm) or Gerris (family Gerridae, popularly known as shoemakers or water skaters), but their size is small, between two and three centimeters in length.
In the cases in which they had more appendages, if it were a decapod crustacean, it would preferably have to be swimming. Currently, some brachyurans, such as Ovalipes trimaculatus that inhabit coastal environments in much of the world (De Grave et al., 2009; Ng et al., 2008), move along the bottom with the first pereiopods or chelipeds (pincers) raised and with the digits of the fifth pereiopods, which are laminar or paddle-shaped, also raised, allowing them to move quickly in semi-swimming, supporting only the three appendages (2nd to 4th pereiopods) on each side. This lateral displacement leaves a set of traces divided into triads, which at first glance could resemble ours. Pasini et al. (2016) described in the Pliocene of the Italian Piedmont tracks left by indeterminate anomuran and brachyuran crabs, showing a rhythmic, laterally asymmetrical series of diagonal steps, parallel to each other or almost superimposed, represented by four pairs of depressed and separated slits, followed by elongated drop-shaped impressions directed laterally. However, in some tracks, only the tips of the three paired gait tracks are preserved, which could resemble our trackway.
The dimensions of this possible crab would not be a problem a priori either, since among the decapod crustaceans there are fossils and current representatives of large size. Feldmann and Schweitzer (2016) describe a giant spider crab (Libinia amplissimus) from the Miocene of the Marys Formation (Maryland, USA) with long and generally highly developed pereiopods (mere and propodia around 20 cm long). And currently, there are some brachyurans such as Pseudocarcinus gigas (Lamarck, 1818) that can have 50 cm long appendages, or the giant Japanese crab or giant spider crab [Macrocheira kaempferi (Temminck, 1836)] which is the largest current arthropod with almost 4 m in length, with front legs that can reach 1.5 m, and a weight that exceeds 20 kg (Clark & Webber, 1991). The problem is that these giant crabs are vagile epibenthic, normally supporting all ten appendages when walking, so they could hardly leave the impression of just three of them. In addition, they inhabit marine environments, some of them (Macrocheira) at a relatively great depth (200 m on average), which is not consistent with the paleoenvironmental context of our site.
On the other hand, the main difference is that the tracks described by Pasini et al. (2016) always present the same orientation, in such a way that the equivalent of our extreme γ always curves to the same side, while, in our case, the orientation of the three parts of the tracks (x, y, z) of the even triads, is opposite to that of the odd ones.
In the case that they were octopods like scorpions, they have three locomotor appendages on each side of the body and a relatively large grasping pincer, while spiders have four locomotor legs. In both cases it seems quite improbable that a lateral transport of the body could leave marks similar to ours, and because, moreover, there is no fossil record of these very large organisms.
Briggs and Rolfe (1983) described a hexapod trail in the Lower Carboniferous of Pennsylvania (USA) whose pattern and, above all, size, could be assimilated a priori to the one studied here. Thus, its trackway presents remarkably constant cross-sectional dimensions, with a total width of 79 cm, formed by a set of tracks made up of three different paired tracks. The shape of all three tracks is concave adaxially, with a pronounced posterior curve or angle halfway along their length. The external ones are the largest, hook-shaped, adaxially concave, with the distal wall more inclined than the proximal one; the intermediate tracks are smaller, sigmoidal, and symmetrical in cross-section and the inner tracks are the smallest. In addition, these sets of tracks are distributed on each side of a shallow median groove, which would imply a continuous displacement supporting all the appendages at the same time. The authors included these tracks in the ichnotaxon Palmichnium Richter, 1954, to differentiate it from Paleohelcura Gilmore, 1926, which is characterized by long parallel and continuous rows of appendage tracks arranged in groups of three, but occasionally with undulations between them and arranged in a straight line inclined at an angle of 60º–80º with respect to the midline.
An ichnotaxon that may have some similarities to a body being transported laterally in a stream is Monomorphichnus Crimes, 1970. However, this consists of a series of straight or slightly sigmoidal striae, usually associated in pairs and which may be repeated laterally. One striation of each pair is usually more prominent than the other and is bifid at one end. On the other hand, the size is also different, as these striae are very shallow and barely exceed 4 cm in length. These characteristics set it apart from those observed here, among other things, because the producer of Monomorphichnus Crimes, 1970, is attributed to the impression by several clawed limbs of trilobites, interpreted as a swimming-grazing trail.
Obviously, all these characteristics separate them from our tracks, so the only thing that can be concluded is that the tracks described here seem to correspond to a large organism (metric scale), which left the impression of three appendages alternately, either for having a hexapod-type organization, or decapod (or even octopod) with particularities and that was dragged laterally by a current in a very shallow lacustrine or wetland environment.
6 Conclusions
The presence of tracks that present a pattern formed by sets of three tracks or triads, which are arranged subparallel to each other, and are distributed in alternating groups, is studied. From a morphological point of view, each track is characterised by a depression formed by a central body three times longer than wide, with straight or slightly curved walls. At each end, there are also two bodies of lesser development, one shorter and more pronounced than the opposite one, which is more elongated. All biomechanical possibilities that could be compatible with an anatomical design that could leave the impression of three alternating appendages are considered. It is concluded that it must have been a giant arthropod with a hexapod or decapod (less likely octopod) organisation. Given its spatial distribution and its relationship with the lithological and sedimentological characteristics, the most likely hypothesis for its formation would be that of an organism swept laterally by a current in a very shallow lacustrine or wetland environment. No fossil organism, nor its present-day equivalent, is known to correspond to these characteristics.
Data availability
The data and matherial supporting their findings can be found from the text citations and localizations.
Change history
19 October 2023
A Correction to this paper has been published: https://doi.org/10.1007/s41513-023-00222-w
References
Baena, J. (1981). Mapa Geológico de España E 1:50.000, 2ª serie. Hoja nº 869 (Jumilla). IGME. Secretaría de Publicaciones, Ministerio de Industria.
Bolton, H. (1911). On a collection of insect-remains from the south wales coalfield. Quarterly Journal of the Geological Society, 67, 149–174. https://doi.org/10.1144/GSL.JGS.1911.067.01-04.06
Bolton, H. (1921). A monograph of the fossil insects of the British coal measures. Part I (pp. 1–80). Paleontographical Society. https://doi.org/10.5962/bhl.title.2477
Briggs, D. E. K., & Rolfe, W. D. I. (1983). A giant arthropod trackway from the Lower Mississipian of Pennsylvania. Journal of Paleontology, 57, 377–390.
Bromley, R. G., Uchman, A., Gregory, M. R., & Martin, A. J. (2003). Hillichnus lobosensis igen. et isp. nov., a complex trace fossil produced by tellinacean bivalves, Paleocene, Monterey, California, USA. Paleogeography, Palaeoclimatology, Palaeoecology, 192, 157–186. https://doi.org/10.1016/S0031-0182(02)00684-3
Brown, T. (1999). The science and art of tracking (p. 219). Berkley Books.
Clark, P., & Webber, W. (1991). A redescription of Macrocheira kaempferi (Temminck, 1836) zoeas with a discussion of the classification of the Majoidea Samouelle, 1819 (Crustacea: Brachyura). Journal of Natural History, 25(5), 1259–1279. https://doi.org/10.1080/00222939100770781
Crimes, T. P. (1970). Trilobite tracks and other trace fossils from the Upper Cambrian of North Wales. Geological Journal, 7, 47–68. https://doi.org/10.1002/gj.3350070104
De Grave, S., Pentcheff, N. D., Ahyong, S. T., Chan, T.-Y., Crandall, K. A., Dworschak, P. C., Felder, D. L., Feldmann, R. M., Fransen, C. H. J. M., Goulding, L. Y. D., Lemaitre, R., Low, M. E. Y., Martin, J. W., Ng, P. K. L., Schweitzer, C. E., Tan, S. H., Tshudy, D., & Wetzer, R. (2009). A classification of living and fossil genera of decapod crustaceans. Raffles Bulletin of Zoology, 21, 1–109.
De Miguel, D., Azanza, B., & Morales, J. (2019). Regional impacts of global climate change: A local humid phase in central Iberia in a late Miocene drying world. Palaeontology, 62, 77–92. https://doi.org/10.1111/pala.12382
Feldmann, R. M., & Schweitzer, C. E. (2016). Giant spider crab from the St. Marys Formation (Miocene) in Calvert County, Maryland, USA. Bulletin of the Mizunami Fossil Museum, 42, 23–28.
Gatesy, S. M. (2003). Direct and indirect tracks feature: What sediment did a dinosaur touch? Ichnos, 10, 91–98. https://doi.org/10.1080/10420940390255484
Getty, P. R., Sproule, R., Stimson, M. R., & Lyons, P. C. (2017). Invertebrate fossils from the Pennsylvanian Rhode Island Formation of Massachusetts, USA. Atlantic Geology, 53, 185–206. https://doi.org/10.4138/atlgeol.2017.007
Gilmore, C. W. (1926). Fossil footprints from the Grand Canyon. Smithsonian Miscellaneous Collections, 77, 1–41.
Herrero, C., Herrero, E., Martín-Chivelet, J., & Pérez-Lorente, F. (2022). Vertebrate ichnofauna from Sierra de las Cabras tracksite (Late Miocene, Jumilla, SE Spain). Mammalian ichnofauna. Journal of Iberian Geology, 48, 241–279. https://doi.org/10.1007/s41513-022-00192-5
Herrero, C., Herrero, E., Martín-Chivelet, J., & Pérez-Lorente, F. (2023). Avian ichnofauna from Sierra de las Cabras tracksite (late Miocene, Jumilla, SE Spain). Journal of Iberian Geology, 49, 31–46. https://doi.org/10.1007/s41513-023-00205-x
Jiménez-Moreno, G., Fauquette, S., & Suc, J. P. (2010). Miocene to Pliocene vegetation reconstruction and climate estimates in the Iberian Peninsula from pollen data. Review of Palaeobotany and Palynology, 162, 403–415. https://doi.org/10.1016/j.revpalbo.2009.08.001
Lamarck, J. B. (1818). Histoire naturelle des Animaux sans Vertèbres. Deterville.
Linnaeus, C. (1758). Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata [10th revised edition] (vol. 1, 824 pp). Laurentius Salvius. https://doi.org/10.5962/bhl.title.542
Martín, J. M., Braga, J. C., Aguirre, J., & Puga-Bernabéu, A. (2009). History and evolution of the North-Betic Strait (Prebetic Zone, Betic Cordillera): A narrow, early Tortonian, tidal-dominated, Atlantic-Mediterranean marine passage. Sedimentary Geology, 216, 80–90. https://doi.org/10.1016/j.sedgeo.2009.01.005
Ng, P. K. L., Guinot, D., & Davie, P. J. F. (2008). Systema Brachyurorum: Part I. An annotated checklist of extant Brachyuran crabs of the world. Raffles Bulletin of Zoology, 17, 1–286.
Pasini, G., Baldanza, A., Gallo, L. M., Garassino, A., & Karasawa, H. (2016). Anomuran and brachyuran trackways and resting trace from the Pliocene of Valduggia (Piedmont, NW Italy): Environmental, behavioural, and taphonomic implications. Natural History Sciences. Atti Della Società Italiana Di Scienze Naturali e Del Museo Civico Di Storia Naturale in Milano, 3, 35–48. https://doi.org/10.4081/nhs.2016.281
Requeta, L. E., Hernández Medrano, N., & Pérez-Lorente, F. (2006–7). La Pellejera: Descripción y aportaciones. Heterocronía y variabilidad de un yacimiento con huellas de dinosaurio de La Rioja (España). Zubía monográfico, 18(1), 21–114.
Richter, R. (1954). Fährte eines ‘Riesenkrebses’ im Rheinischen Schiefergebirge. Natur Und Volk, 84, 261–269.
Roca, E., Sans, M., & Koyi, H. (2006). Polyphase deformation of diapiric areas in models and in the eastern Prebetics (Spain). American Association of Petroleum Geologists Bulletin, 90, 115–136.
Rossi, C., Vilas, L., & Arias, C. (2015). The Messinian marine to nonmarine gypsums of Jumilla (Northern Betic Cordillera, SE Spain): Isotopic and Sr concentration constraints on the origin of parent brines. Sedimentary Geology, 328, 96–114. https://doi.org/10.1016/j.sedgeo.2015.08.007
Rubinat, M., Roca, E., Escalas, M., Queralt, P., Ferrer, O., & Ledo, J. J. (2013). The influence of basement structure on the evolution of the Bicorb-Quesa Diapir (eastern Betics Iberian Peninsula): Contractive thin-skinned deformation above a pre-existing extensional basement fault. International Journal of Earth Sciences, 102, 25–41. https://doi.org/10.1007/s00531-012-0789-9
Sarjeant, W. A. S. (1989). “Ten paleoichnological commandments”: A standardized procedure for the description of fossil vertebrate footprints. In D. D. Gillette & M. G. Lockley (Eds.), Dinosaur Tracks and Traces (pp. 369–370). Cambridge Univ.
Temminck, C. J. (1836). Coup-d'oeil sur la faune des Iles de la Sonde et de l'Empire du Japon. Discours préliminaire destiné a servir d'introduction a la Faune du Japon. pp. 30. https://doi.org/10.5962/bhl.title.119899
Thulborn, R. A., & Wade, M. (1989). A footprint as a history of movement. In D. D. Gillette & M. G. Lockley (Eds.), En Dinosaurs Tracks and Traces (pp. 51–56). Cambridge Univ Press.
van Dam, J. A. (2006). Geographic and temporal patterns in the late Neogene (12–3 Ma) aridification of Europe: The use of small mammals as paleoprecipitation proxies. Palaeogeography, Palaeoclimatology, Palaeoecology, 238, 190–218. https://doi.org/10.1016/j.palaeo.2006.03.025
Walker, F. (1871). Catalogue of the Specimens of Dermaptera saltatoria in the Collection of the British Museum. Part V. Supplement to the Catalogue of Blattariae, British Museum (Natural History) London. British Museum (Natural History), London 40.
Wood, G. (1983). The Guinness Book of Animal Facts and Feats. Sterling Pub Co Inc; Subsequent edition (December 31, 1899).
Acknowledgements
Thanks to Nicholas Minter an anonymous reviewer for comments and suggestions, which improved the manuscript. This study has been partially supported by the project PID2019-104625RB-100 of the Ministry of Science and Innovation, State Research Agency, and by the Junta de Andalucía to the Research Group RNM276 of the CCTH of the University of Huelva.
Funding
Funding for open access publishing: Universidad de Huelva/CBUA. Funding for open access charge: Universidad de Huelva/CBUA.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
The original online version of this article was revised: The name assigned to the trace fossil Aenigmatichnus has already been used in another publication. It was therefore changed to Aenigmatipodus in this paper.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mayoral, E., Herrero, C., Herrero, E. et al. A new enigmatic lacustrine trackway in the upper Miocene of the Sierra de las Cabras (Jumilla, Murcia, Spain). J Iber Geol 49, 237–256 (2023). https://doi.org/10.1007/s41513-023-00215-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-023-00215-9
Keywords
- Aenigmatipodus jumillensis nov. ichnogen. nov. ichnosp.
- Arthropod trace
- Lacustrine environment
- Southeast Iberia
- Upper Miocene